原厂验证网址:https://xonamicrofluidics.com/contact/
美国XonaMicrofluidics公司神经元突触轴突培养板 现货供应
型号:内详
价格:请致电:010-67529703
品牌:xonamicrofluidics
美国XonaMicrofluidics公司神经元突触轴突培养板 现货供应

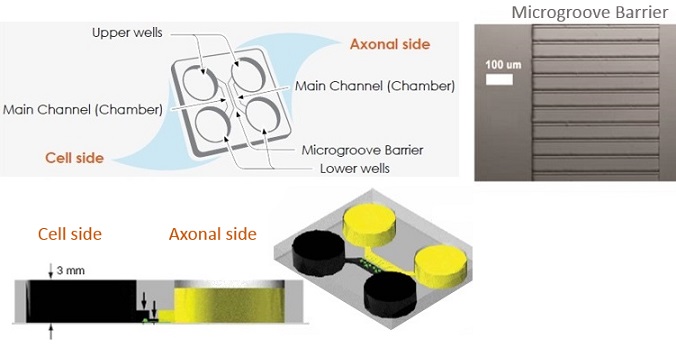
XONA MICROFLUIDICS提供了多种用于神经元细胞培养的设备,包括:
SILICONE DEVICES神经元突触轴突连接微流体培养板

美国Xona Microfluidics LLC公司提供的神经元装置是光学透明的聚二甲基硅氧烷(PDMS)和生物惰性制造而成。神经元装置由微槽连接的两个腔室组成。神经元装置可以使研究者通过微流体方式隔离细胞体与轴突。
Xona提供多种硅胶设备配置。 SND是原始的2隔室。 TCND具有3个隔室,可提供多种实验可能性。 圆形设备(RD)与精选的玻璃底皿兼容,使其可用于活细胞成像。
我们锁有的硅胶(PDMS)腔室都需要终用户自由组装。
XonaChips®是神经元培养的新创新!
XonaChips®已预先组装和预先粘合,比我们原始的有机硅设备更易于使用。 这些塑料芯片可改善动物模型神经元的长期培养。 它们对于培养人干细胞来源的神经元te别有用,可提供更好的细胞附着和长期生长。
XonaChips®是光学透明的,非常适合荧光实时成像。

目前,Xona正在提供两室XonaChips®XC系列,具有150、450或900 µm的微槽屏障。 每盒(5)XC系列XonaChips®都装有2 ml XC Pre-Coat™管。
目前,Xona还提供带有3个隔室的XonaChips®XC-T系列,该隔室带有(2)个500 µm的微沟槽屏障,其中有500 um的中央隔室或1000 um的中央隔室。 每盒(5)XC-T系列XonaChips®包括2(2)ml XC Pre-Coat™管。
XC Pre-Coat™和XonaPDL™经过化,可与XonaChips®一起使用。
ChipTray™

长期培养神经元过程中培养基的蒸发对细胞健康有很大影响。 在具有较小孔的微流体装置和芯片中尤其如此。 为了克服这些问题,Xona开发了一种获得专利的ChipTray™,可与XonaChips®,我们的有机硅设备以及锁有其他微流体设备一起使用。 除了减少蒸发之外,ChipTray™还提供了化的外壳,用于使用空气物镜进行相衬成像。
托盘的尺寸与标准ANSI微孔板支架兼容。 托盘的两端有两个水槽,两个用于XonaChips®的槽(XC150,XC450,XC900,XC-T-500或XC-T-1000)。 安装在玻璃上的有机硅设备或您要使用的任何其他基于幻灯片的微流体设备也与这些托盘兼容。 在每个插槽中都有一个切口,可使用倒置显微镜提高图像质量。 ChipTrays已准备好进行组织培养。
|
订货号 |
简述 |
货物状态 |
| xona chip | ||
|
XC150 |
XonaChip 150 um barrier 5PK includes 2 ml tube XC Pre-Coat |
现货促销 |
|
XC450 |
XonaChip 450 um barrier 5PK includes 2 ml tube XC Pre-Coat |
现货促销 |
|
XC900 |
XonaChip 900 um barrier 5PK includes 2 ml tube XC Pre-Coat |
现货促销 |
|
XC-T500 |
XonaChip 500 um central compartment includes (2) 2 ml tubes XC Pre-Coat |
现货促销 |
|
XC-T1000 |
XonaChip 1000 um central compartment includes (2) 2 ml tubes XC Pre-Coat |
现货促销 |
|
XonaPDL |
Optimized poly-d-lysine solution for Xona platforms |
现货促销 |
| SILICONE DEVICES ROUND (RD) | ||
|
RD150 |
Round Device 150 um barrier 5PK |
现货促销 |
|
RD450 |
Round Device 450 um barrier 5PK |
现货促销 |
|
RD900 |
Round Device 900 um barrier 5PK |
现货促销 |
| SILICONE DEVICES 2-COMPARTMENT (SND) | ||
|
SND150 |
Standard Neuron Device 150 um barrier 5PK |
现货促销 |
|
SND450 |
Standard Neuron Device 450 um barrier 5PK |
现货促销 |
|
SND900 |
Standard Neuron Device 900 um barrier 5PK |
现货促销 |
| SILICONE DEVICES 3-COMPARTMENT (TCND) | ||
|
TCND500 |
Triple Compartment 500 um central compartment 5PK |
现货促销 |
|
TCND1000 |
Triple Compartment 1000 um central compartment 5PK |
现货促销 |
|
RD75 |
Round Device 75 um barrier 5PK |
现货促销 |
|
SND75 |
Standard Neuron Device 75 um barrier 5PK |
现货促销 |
|
uLP |
Microfluidic Local Perfusion Device 5PK |
现货促销 |
|
ChipTray™ (pack of 6) |
Pack of (6) |
现货促销 |
|
ChipTray™ (1 each) |
Pack of 1 |
现货促销 |
|
产品名称 |
产品描述 |
|
RD150 神经元突触细胞微灌流系统 |
圆形设备(150微米的微槽屏障) |
|
RD450 神经元突触细胞微灌流 |
Round Device (450 um microgroove barrier) |
|
RD900 神经元突触细胞微灌流 |
Round Device (900 um microgroove barrier) |
|
SND150 神经元突触细胞微灌流 |
Standard Neuron Device (150 um microgroove
barrier) |
|
SND450 神经元突触细胞微灌流 |
Standard Neuron Device (450 um microgroove
barrier) |
|
SND900 神经元突触细胞微灌流 |
Standard Neuron Device (900 um microgroove
barrier) |
|
TCND500 500um三腔神经元设备 |
三腔神经元设备(500 um微槽屏障) |
|
TCND1000 1000um三腔神经元设备 |
三腔神经元设备(1000 um
微槽屏障) |
|
uLP |
>微流局部灌流装置
屏障,(50 um microgroove 75微米灌注通道,500 um microgroove
障碍) |
2018
Exosomes derived from high-glucose-stimulated Schwann cells promote development of diabetic peripheral neuropathy.
FASEB J. 2018 Jun 22:fj201800597R. doi: 10.1096/fj.201800597R. [Epub ahead of print]
Microfluidics of Small-Population Neurons Allows for a Precise Quantification of the Peripheral Axonal Growth State
Front Cell Neurosci. 2018; 12: 166. June 15, 2018
Advances in ex vivo models and lab-on-a-chip devices for neural tissue engineering.
Biomaterials. 2018 May 11. pii: S0142-9612(18)30347-8. doi: 10.1016/j.biomaterials.2018.05.012. [Epub ahead of print]
Neuronal subtype determines HSV-1 Latency-Associated-Transcript (LAT) promoter activity during latency.
J Virol. 2018 Apr 11. pii: JVI.00430-18. doi: 10.1128/JVI.00430-18. [Epub ahead of print]
Axonal Activation of the Unfolded Protein Response Promotes Axonal Regeneration Following Peripheral Nerve Injury.
Neuroscience. 2018 Apr 1;375:34-48. doi: 10.1016/j.neuroscience.2018.02.003. Epub 2018 Feb 10.
Exosomes taken up by neurons hijack the endosomal pathway to spread to interconnected neurons.
Acta Neuropathol Commun. 2018 Feb 15;6(1):10. doi: 10.1186/s40478-018-0514-4.
Neurturin is a PGC-1a1-controlled myokine that promotes motor neuron recruitment and neuromuscular junction formation.
Mol Metab. 2018 Jan;7:12-22. doi: 10.1016/j.molmet.2017.11.001. Epub 2017 Nov 7.
Importin α1 is required for nuclear import of herpes simplex virus proteins and capsid assembly in fibroblasts and neurons.
PLoS Pathog. 2018 Jan 5;14(1):e1006823. doi: 10.1371/journal.ppat.1006823. eCollection 2018 Jan.
Visualization of local phosphatidylcholine synthesis within hippocampal neurons using a compartmentalized culture system and imaging mass spectrometry
Biochem Biophys Res Commun. 2018 Jan 1;495(1):1048-1054. doi: 10.1016/j.bbrc.2017.11.108. Epub 2017 Nov 21.
Intercellular transfer of pathogenic a-synuclein by extracellular vesicles is induced by the lipid peroxidation product 4-hydroxynonenal
Neurobiol Aging. 2018 Jan;61:52-65. doi: 10.1016/j.neurobiolaging.2017.09.016. Epub 2017 Sep 22.
2017
Distal axotomy enhances retrograde presynaptic excitability onto injured pyramidal neurons via trans-synaptic signaling.
Nat Commun. 2017 Sep 20;8(1):625.
Microfluidic Device for Studying Traumatic Brain Injury
Part of the Neuromethods book series (NM, volume 126) First Online: 12 May 2017
Mild and repetitive very mild axonal stretch injury triggers cystoletal mislocalization and growth cone collapse
PLoS One. 2017; 12(5): e0176997. Published online 2017 May 4.
Using high-throughput barcode sequencing to efficiently map connectomes
Nucleic Acids Res. 2017 Jul 7. 45(12): e115. Published online 2017 Apr 26
Messenger RNAs localized to distal projections of human stem cell derived neurons.
Sci Rep 7. 2017 Apr 4;7(1):611. Published online
NGF-dependent axon growth and regeneration are altered in sympathetic neurons of dystrophic mdx mice.
Mol Cell Neurosci. 2017 Feb 2;80:1-17 [Epub ahead of print]
T-type Ca2+ channels are required for enhanced sympathetic axon growth by TNFα reverse signaling Open Biology. Published 18 January 2017.DOI: 10.1098/rsob.160288
The release and trans-synaptic transmission of Tau via exosomes
Mol Neurodegener. 2017; 12: 5. Published online 2017 Jan 13.
MiR-29c/PRKCI Regulates Axonal Growth of Dorsal Root Ganglia Neurons Under Hyperglycemia.
Mol Neurobiol. 2017 Jan 9. [Epub ahead of print]
Co-culture Synaptogenic Assay: A New Look at Fluorescence Reporters and Technological Devices.
Methods Mol Biol. 2017;1538:13-27qua
2016
Acetylated tubulin is essential for touch sensation in mice
eLife. 2016; 5: e20813. Published online 2016 Dec 13
Mesenchymal Stromal Cells Promote Axonal Outgrowth Alone and Synergistically with Astrocytes via tPA
PLoS One. 2016; 11(12): e0168345. Published online 2016 Dec 13.
Compartmentalized Microfluidic Platforms: The Unrivaled Breakthrough of In Vitro Tools for Neurobiological Research.
J Neurosci. 2016 Nov 16;36(46):11573-11584.
Pathological α-synuclein transmission initiated by binding lymphocyte-activation gene 3
Science. 2016 Sep 30; 353(6307): aah3374.
TRiC subunits enhance BDNF axonal transport and rescue striatal atrophy in Huntington’s disease
Proc Natl Acad Sci U S A. 2016 Sep 20; 113(38): E5655–E5664. Published online 2016 Sep 6.
Interneuronal Transfer and Distal Action of Tetanus Toxin and Botulinum Neurotoxins A and D in Central Neurons.
Cell Rep. 2016 Aug 16;16(7):1974-87. doi: 10.1016/j.celrep.2016.06.104. Epub 2016 Aug 4.
Probing extracellular Sonic hedgehog in neurons.
Biol Open. 2016 Aug 15;5(8):1086-92. doi: 10.1242/bio.019422.
Leucine Zipper-bearing Kinase promotes axon growth in mammalian central nervous system neurons
Sci Rep. 2016; 6: 31482. Published online 2016 Aug 11.
miR-145 Regulates Diabetes-Bone Marrow Stromal Cell-Induced Neurorestorative Effects in Diabetes Stroke Rats.
Stem Cells Transl Med. 2016 Jul 26. pii: sctm.2015-0349. [Epub ahead of print]
Semaphorin 3C Released from a Biocompatible Hydrogel Guides and Promotes Axonal Growth of Rodent and Human Dopaminergic Neurons.
Tissue Eng Part A. 2016 Jun;22(11-12):850-61. doi: 10.1089/ten.TEA.2016.0008.
A New Defective Helper RNA to Produce Recombinant Sindbis Virus that Infects Neurons but does not Propagate
Front Neuroanat. 2016; 10: 56. Published online 2016 May 24. doi: 10.3389/fnana.2016.00056
Monitoring mRNA Translation in Neuronal Processes Using Fluorescent Non-Canonical Amino Acid Tagging.
J Histochem Cytochem. 2016 May;64(5):323-33. doi: 10.1369/0022155416641604. Epub 2016 Mar 29.
MicroRNA 146a locally mediates distal axonal growth of dorsal root ganglia neurons under high glucose and sildenafil conditions.
Neuroscience. 2016 Aug 4;329:43-53. doi: 10.1016/j.neuroscience.2016.05.005. Epub 2016 May 7.
Retrograde fibroblast growth factor 22 (FGF22) signaling regulates insulin-like growth factor 2 (IGF2) expression for activity-dependent synapse stabilization in the mammalian brain.
Elife. 2016 Apr 15;5. pii: e12151. doi: 10.7554/eLife.12151.
Exosomes Derived from Mesenchymal Stromal Cells Promote Axonal Growth of Cortical Neurons.
Mol Neurobiol. 2016 Mar 19. [Epub ahead of print]
The RNA-binding protein SFPQ orchestrates an RNA regulon to promote axon viability.
Nat Neurosci. 2016 May;19(5):690-6. doi: 10.1038/nn.4280. Epub 2016 Mar 28.
Quantitative and semi-quantitative measurements of axonal degeneration in tissue and primary neuron cultures
J Neurosci Methods. 2016 Jun 15; 266: 32–41. Published online 2016 Mar 28.
Functional Impact of Corticotropin-Releasing Factor Exposure on Tau Phosphorylation and Axon Transport.
PLoS One. 2016 Jan 20;11(1):e0147250. doi: 10.1371/journal.pone.0147250. eCollection 2016.
Compartmentalized Platforms for Neuro-Pharmacological Research.
Curr Neuropharmacol. 2016;14(1):72-86.
D-4F Decreases White Matter Damage After Stroke in Mice
Stroke. 2016 Jan;47(1):214-20. doi: 10.1161/STROKEAHA.115.011046. Epub 2015 Nov 24.
2015
Dual Role of Herpes Simplex Virus 1 pUS9 in Virus Anterograde Axonal Transport and Final Assembly in Growth Cones in Distal Axons.
J Virol. 2015 Dec 23;90(5):2653-63. doi: 10.1128/JVI.03023-15.
The Basic Domain of Herpes Simplex Virus 1 pUS9 Recruits Kinesin-1 To Facilitate Egress from Neurons.
J Virol. 2015 Dec 9;90(4):2102-11. doi: 10.1128/JVI.03041-15.
Dendrosomatic Sonic Hedgehog Signaling in Hippocampal Neurons Regulates Axon Elongation
J Neurosci. 2015 Dec 9; 35(49): 16126–16141.
MicroRNAs in the axon locally mediate the effects of chondroitin sulfate proteoglycans and cGMP on axonal growth
Dev Neurobiol. 2015 Dec; 75(12): 1402–1419. Published online 2015 Jun 24.
Whole transcriptome profiling reveals the RNA content of motor axons
Nucleic Acids Res. 2016 Feb 29; 44(4): e33. Published online 2015 Oct 12.
Synaptic Contacts Enhance Cell-to-Cell Tau Pathology Propagation. Cell Rep. 2015 May 26;11(8):1176-83
Morphometric assessment of toxicant induced neuronal degeneration in full and restricted contact co-cultures of embryonic cortical rat neurons and astrocytes: using m-Dinitrobezene as a model neurotoxicant. Toxicol In Vitro. 2015 Apr;29(3):564-74.
Coupled local translation and degradation regulate growth cone collapse
Nat Commun. 2015; 6: 6888. Published online 2015 Apr 22.
Control of autophagosome axonal retrograde flux by presynaptic activity unveiled using botulinum neurotoxin type a J Neurosci. 2015 Apr 15;35(15):6179-94
Activation of 5-HT7 receptor stimulates neurite elongation through mTOR, Cdc42 and actin filaments dynamics. Front Behav Neurosci. 2015 Mar 11;9:62.
Identifying the primary site of pathogenesis in amyotrophic lateral sclerosis – vulnerability of lower motor neurons to proximal excitotoxicity
Dis Model Mech. 2015 Mar; 8(3): 215–224.
Recording large extracellular spikes in microchannels along many axonal sites from individual neurons. PLoS One. 2015 Mar 3;10(3):e0118514.
Increased migration of olfactory ensheathing cells secreting the Nogo receptor ectodomain over inhibitory substrates and lesioned spinal cord. Cell Mol Life Sci. 2015 Feb 24.
A ?-secretase inhibitor, but not a ?-secretase modulator, induced defects in BDNF axonal trafficking and signaling: evidence for a role for APP. PLoS One. 2015 Feb 24;10(2):e0118379.
2014
Herpes simplex virus gE/gI extracellular domains promote axonal transport and spread from neurons to epithelial cells. J Virol. 2014 Oct;88(19):11178-86.
PTEN inhibition enhances neurite outgrowth in human embryonic stem cell-derived neuronal progenitor cells. J Comp Neurol. 2014 Aug 15;522(12):2741-55.
A-synuclein immunotherapy blocks uptake and templated propagation of misfolded a-synuclein and neurodegeneration. Cell Rep. 2014 Jun 26;7(6):2054-65.
Regulation of axon growth by the JIP1-AKT axis. Dajas-Bailador F, Bantounas I, Jones EV, Whitmarsh AJ. J Cell Sci. 2014 Jan 1;127(Pt 1):230-9.
2013
Probing functional properties of nociceptive axons using a microfluidic culture system. PLoS One. 2013 Nov 20;8(11)
Microfluidic primary culture model of the lower motor neuron-neuromuscular junction circuit. J Neurosci Methods. 2013 Sep 15;218(2):164-9.
Dendritic GluN2A synthesis mediates activity-induced NMDA receptor insertion. J Neurosci. 2013 May 15;33(20):8898-908.
ProNGF promotes neurite growth from a subset of NGF-dependent neurons by a p75NTR-dependent mechanism. Development. 2013 May;140(10):2108-17.
The MicroRNA-17-92 cluster enhances axonal outgrowth in embryonic cortical neurons. J Neurosci. 2013 Apr 17;33(16):6885-94.
Axonal translation of β-catenin regulates synaptic vesicle dynamics. J Neurosci. 2013 Mar 27;33(13):5584-9.
Target-derived neurotrophins coordinate transcription and transport of bclw to prevent axonal degeneration. J Neurosci. 2013 Mar 20;33(12):5195-207.
Herpes simplex virus membrane proteins gE/gI and US9 act cooperatively to promote transport of capsids and glycoproteins from neuron cell bodies into initial axon segments. J Virol. 2013 Jan;87(1):403-14
Small misfolded Tau species are internalized via bulk endocytosis and anterogradely and retrogradely transported in neurons. J Biol Chem. 2013 Jan 18;288(3):1856-70.
2012
Neuron-to-neuron transmission of α-synuclein fibrils through axonal transport. Ann Neurol. 2012 Oct;72(4):517-24.
Axonal outgrowth and dendritic plasticity in the cortical peri-infarct area after experimental stroke. Stroke. 2012 Aug;43(8):2221-8.
A herpes simplex virus 1 (McKrae) mutant lacking the glycoprotein K gene is unable to infect via neuronal axons and egress from neuronal cell bodies. MBio. 2012 Jul 24;3(4):e00144-12.
Control of viral latency in neurons by axonal mTOR signaling and the 4E-BP translation repressor. Genes Dev. 2012 Jul 15;26(14):1527-32.
Assembly and maintenance of nodes of ranvier rely on distinct sources of proteins and targeting mechanisms. Neuron. 2012 Jan 12;73(1):92-107.
Chronic excitotoxin-induced axon degeneration in a compartmented neuronal culture model. ASN Neuro. 2012 Feb 23;4(1).
Chemical genetic-mediated spatial regulation of protein expression in neurons reveals an axonal function for wld(s). Chem Biol. 2012 Feb 24;19(2):179-87.
Microfluidics Platforms for Investigating Injury and Regeneration of CNS Axons. Ann Biomed Eng. 2012 Jun;40(6):1268-76.
2011
Microfluidic and Compartmentalized platforms for neurobiological research. Crit Rev Biomed Eng. 2011;39(3):185-200.
Neurotrophin-mediated dendrite-to-nucleus signaling revealed by microfluidic compartmentalization of dendrites. Proc Natl Acad Sci U S A. 2011 Jul 5;108(27):11246-51.
Lanosterol induces mitochondrial uncoupling and protects dopaminergic neurons from cell death in a model for Parkinson’s disease. Cell Death Differ. 2012 Mar;19(3):416-27.
The bovine herpesvirus type 1 envelope protein Us9 acidic domain is crucial for anterograde axonal transport. Vet Microbiol. 2011 Sep 28;152(3-4):270-9.
IL-1β inhibits axonal growth of developing sympathetic neurons. Mol Cell Neurosci. 2011 Oct;48(2):142-50.
Micro-scale and microfluidic devices for neurobiology. Curr Opin Neurobiol. 2010 Oct;20(5):640-7.
Examination of axonal injury and regeneration in micropatterned neuronal culture using pulsed laser microbeam dissection. Lab Chip. 2010 Aug 21;10(16):2083-92.
Anterograde transport of herpes simplex virus capsids in neurons by both Separate and Married mechanisms. J Virol. 2011 Jun;85(12):5919-28.
Transcriptome analysis of embryonic and adult sensory axons reveals changes in mRNA repertoire localization. RNA. 2011 Jan;17(1):85-98.
Regulation of trafficking of activated TrkA is critical for NGF-mediated functions. Traffic. 2011 Apr;12(4):521-34.
2010
Microfluidic local perfusion chambers for the visualization and manipulation of synapses. Neuron. 2010 Apr 15;66(1):57-68.
2009
Shear stress effect on transfection of neurons cultured in microfluidic devices. J Nanosci Nanotechnol. 2009 Dec;9(12):7330-5.
Novel microfluidic platform for culturing neurons: culturing and biochemical analysis of neuronal components. Biotechnol J. 2009 Nov;4(11):1573-7.
Axonal elongation triggered by stimulus-induced local translation of a polarity complex protein. Nat Cell Biol. 2009 Aug;11(8):1024-30.
beta-Amyloid impairs axonal BDNF retrograde trafficking. Neurobiol Aging. 2011 May;32(5):821-33.
Axonal mRNA in uninjured and regenerating cortical mammalian axons. J Neurosci. 2009 Apr 15;29(15):4697-707.
Presynaptic regulation of astroglial excitatory neurotransmitter transporter GLT1. Neuron. 2009 Mar 26;61(6):880-94.
2008
A microfluidic chamber for analysis of neuron-to-cell spread and axonal transport of an alpha-herpesvirus. PLoS One. 2008 Jun 18;3(6):e2382.
2006
Microfluidic chambers for cell migration and neuroscience research. Methods Mol Biol. 2006;321:167-77.
Microfluidic culture platform for neuroscience research. Nat Protoc. 2006;1(4):2128-36.
2005
A microfluidic culture platform for CNS axonal injury, regeneration and transport. Nat Methods. 2005 Aug;2(8):599-605.
世联博研北京公司作为美国xonamicrofluidics代商,提醒注意假冒产品,耽误研究
原厂验证网址:https://xonamicrofluidics.com/contact/



